Water pools and species interactions
Miquel De Caceres (CREAF)
2026-07-30
Source:vignettes/workedexamples/PlantWaterPools.Rmd
PlantWaterPools.RmdAbout this vignette
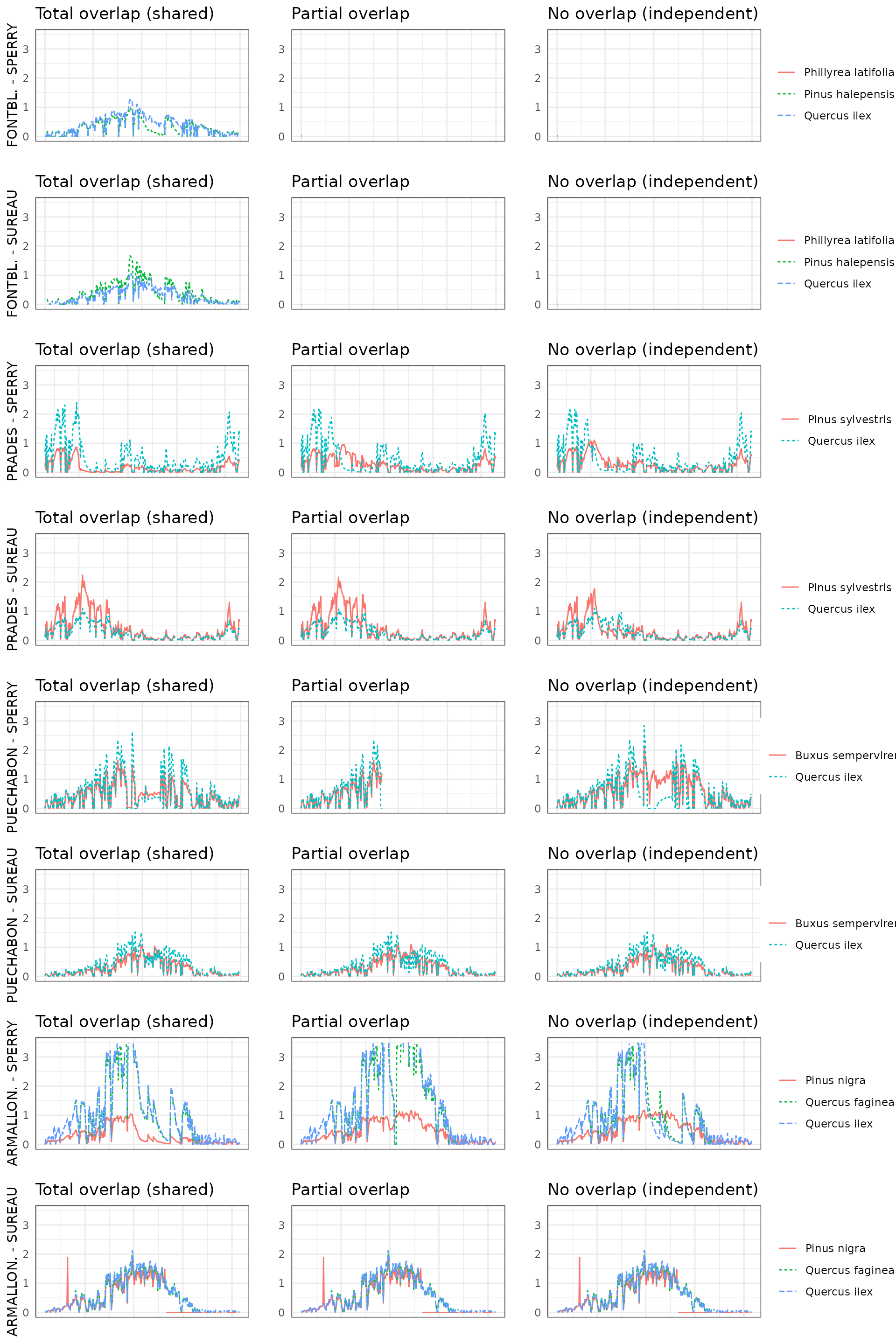
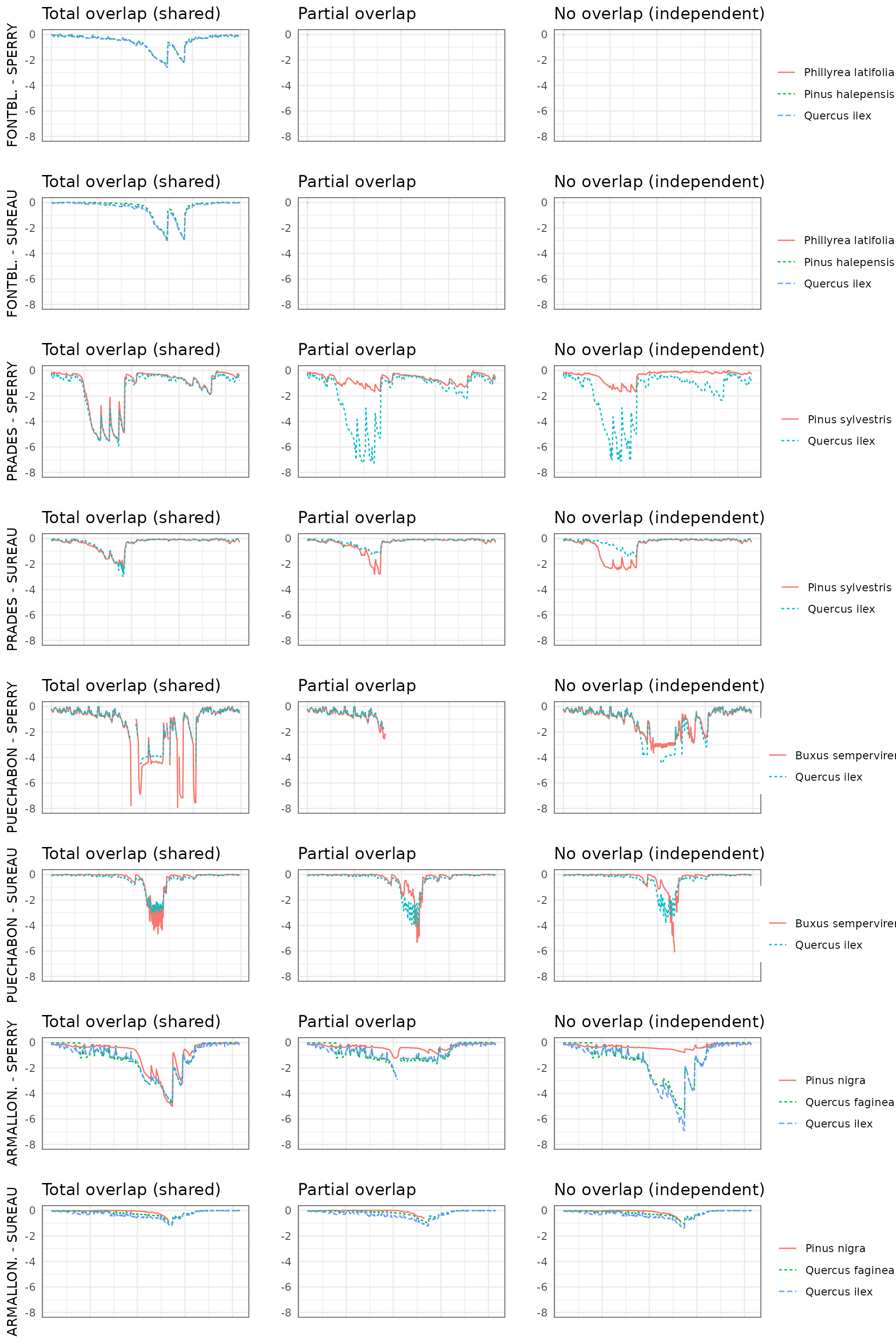
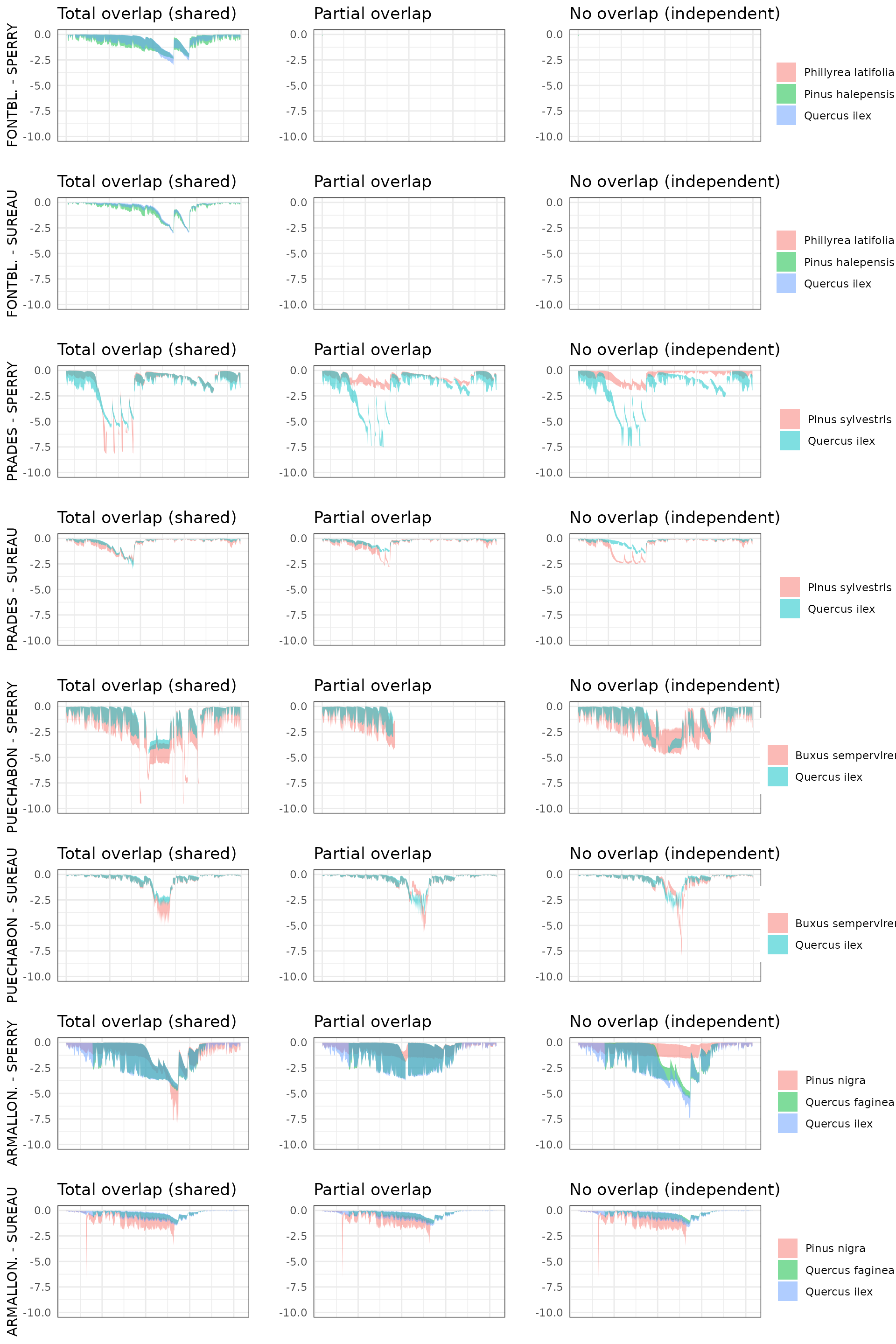
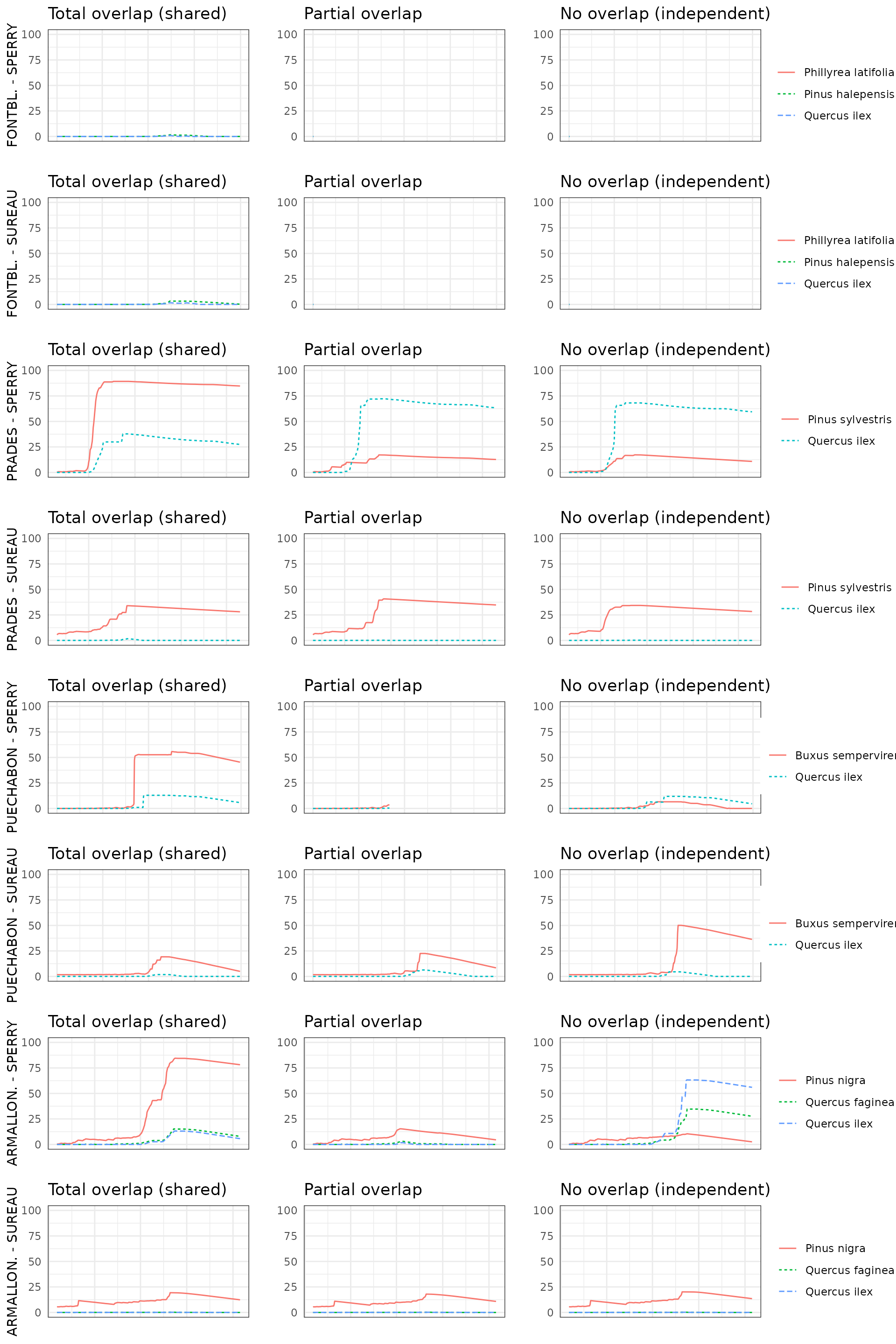
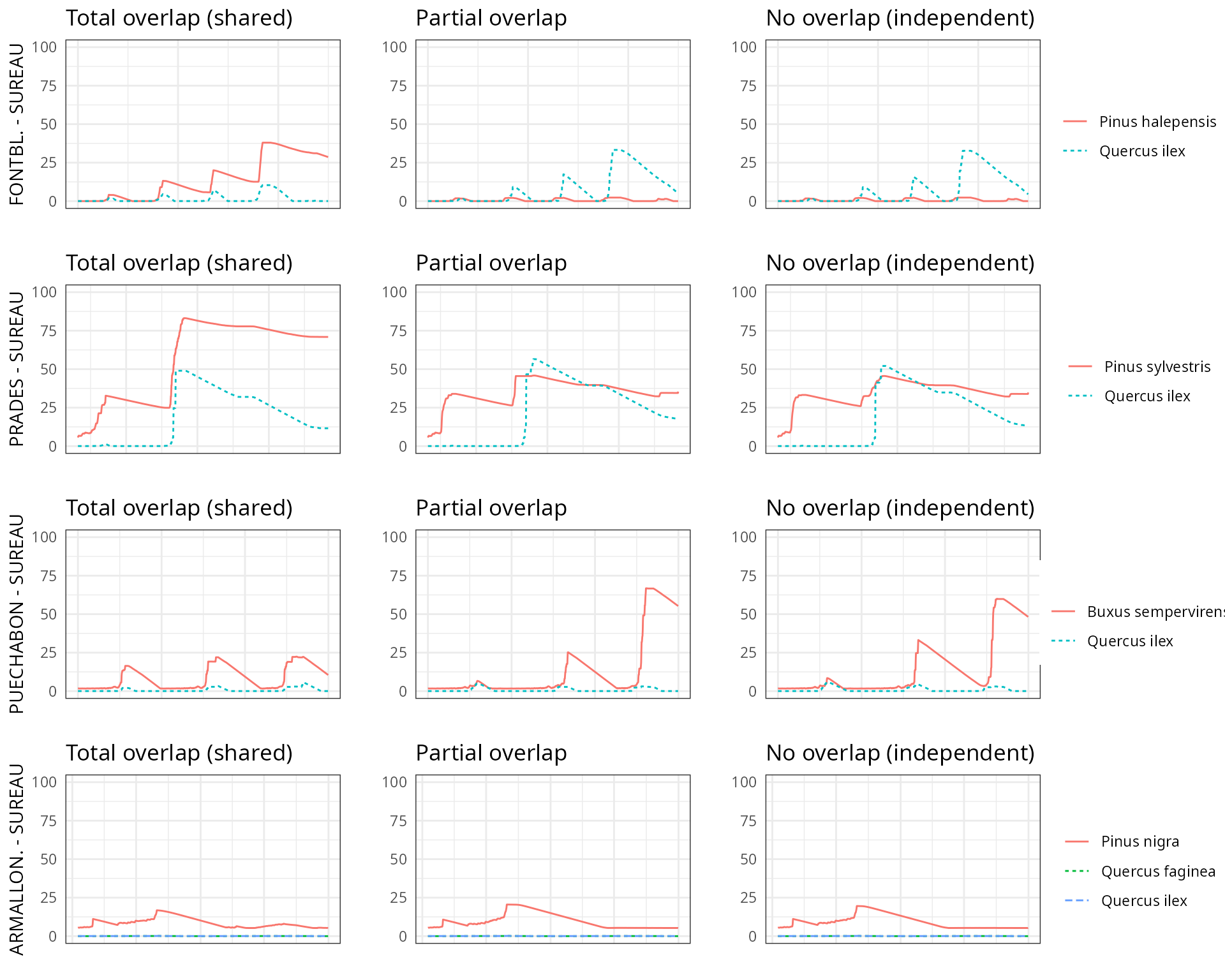
By default, simulations in medfate assume that soil
moisture under all plant cohorts is the same (i.e. water sources
corresponding to vertical soil layers are shared among cohorts).
Therefore, the models neglect spatial variation in soil moisture
observed in mixed forest stands (Schume et al. 2004). Variations in soil
moisture beneath plant cohorts (and, implicitly, horizontal variation of
soil moisture) can be simulated in medfate if required
by the user (see control parameter rhizosphereOverlap).
This involves considering that a given plant cohort will perform water
uptake from the water pool surrounding its roots,
whereas it may not have access to the water beneath other plants.
However, there can exist some degree of horizontal overlap between water
pools exploited by different plants.
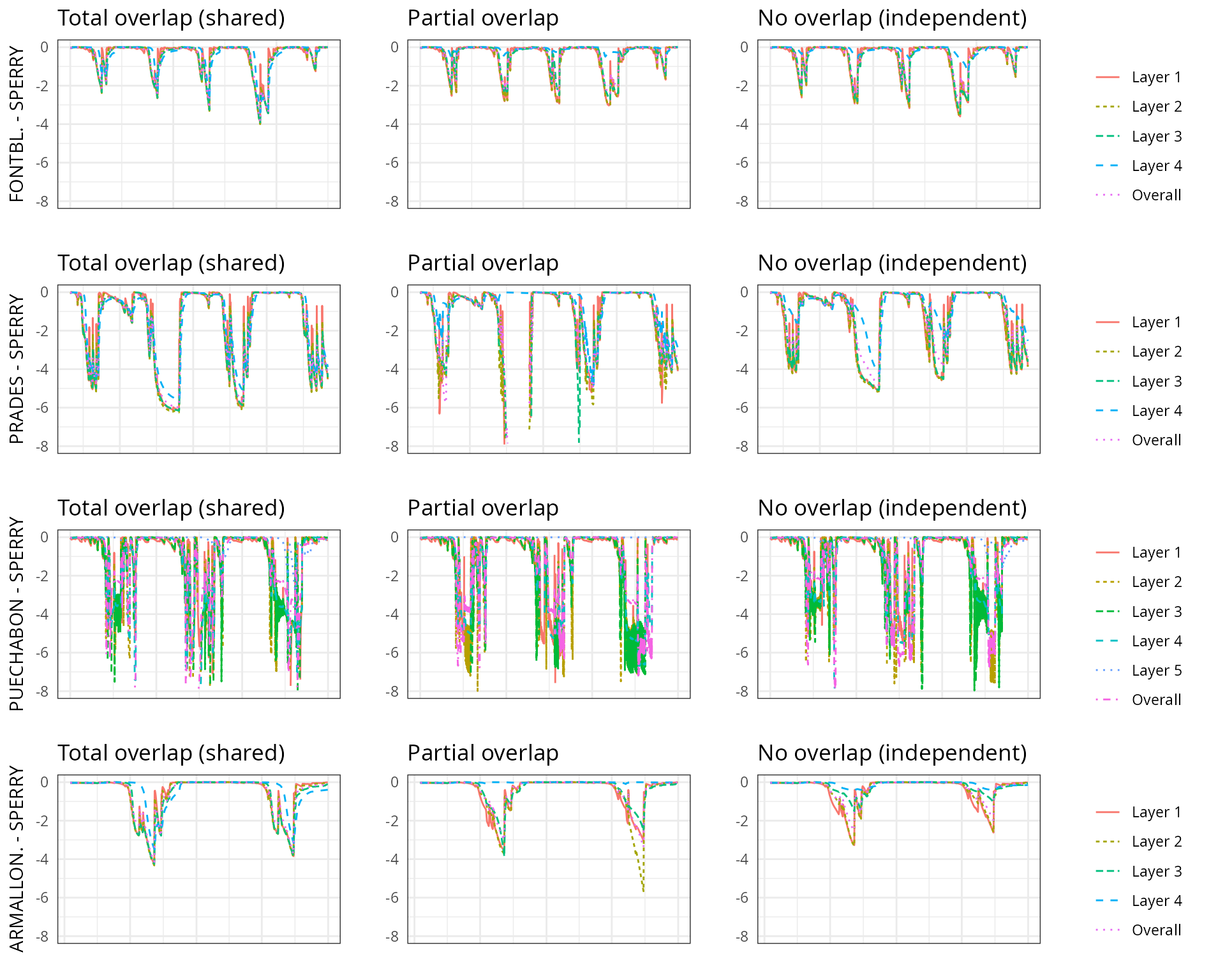
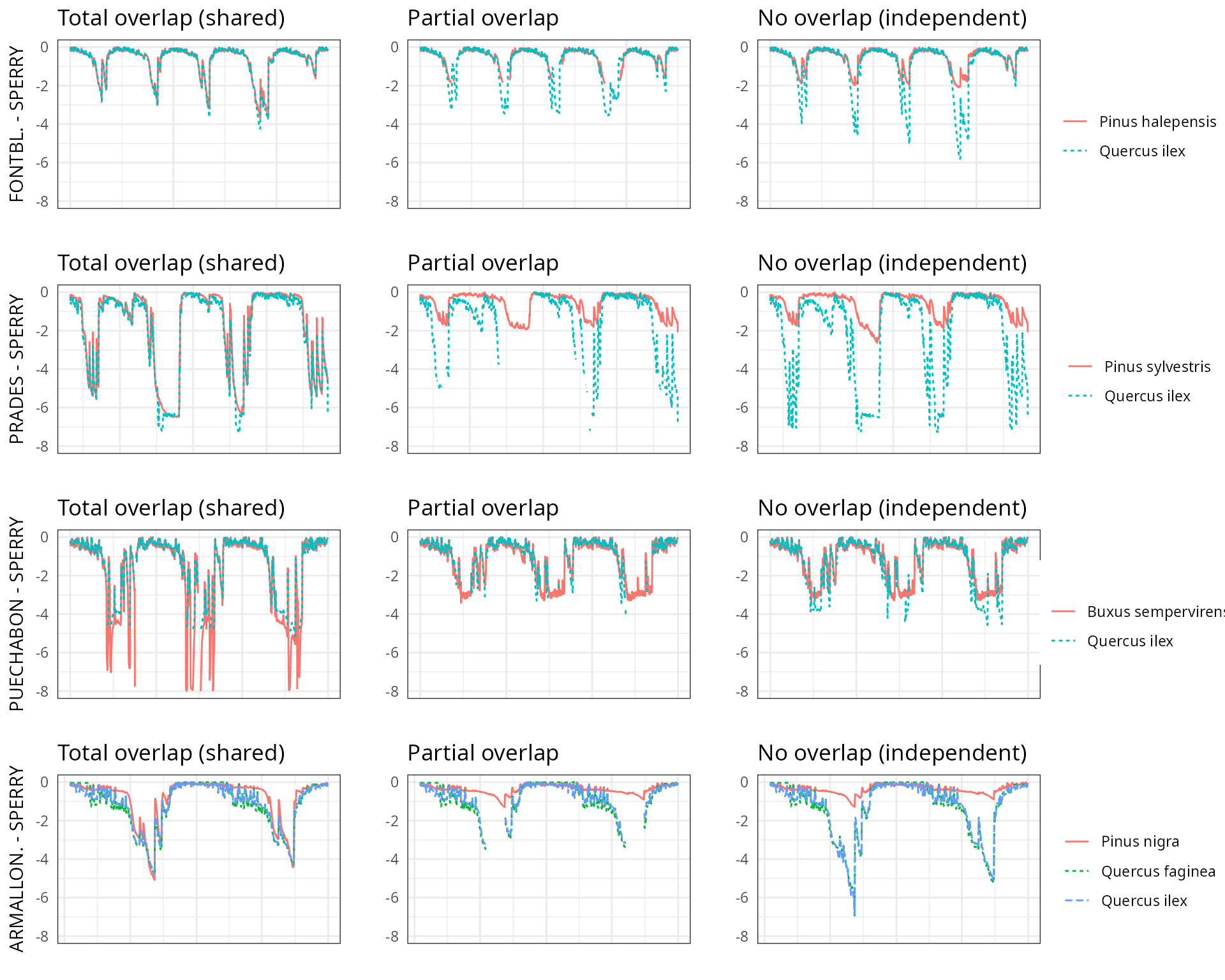
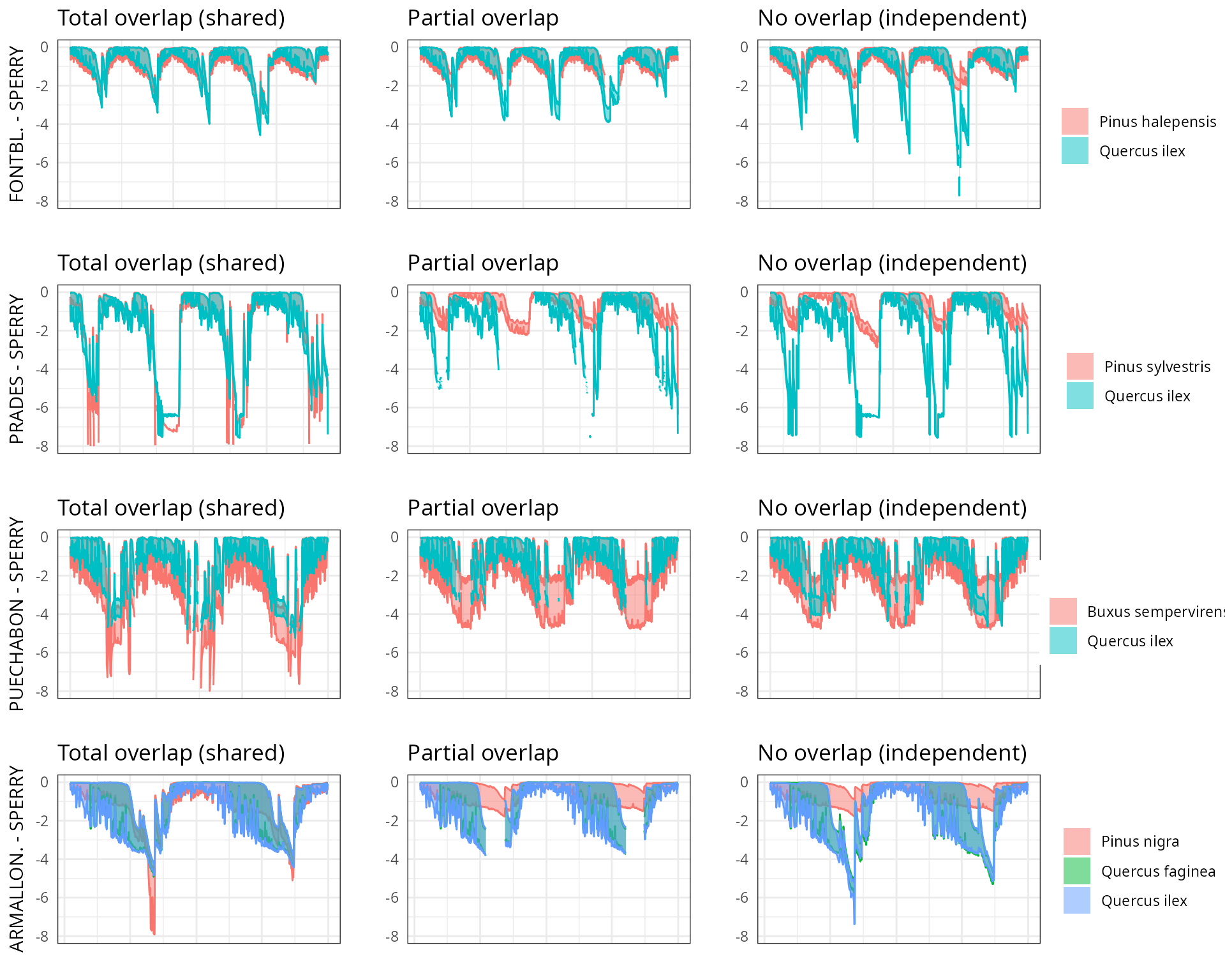
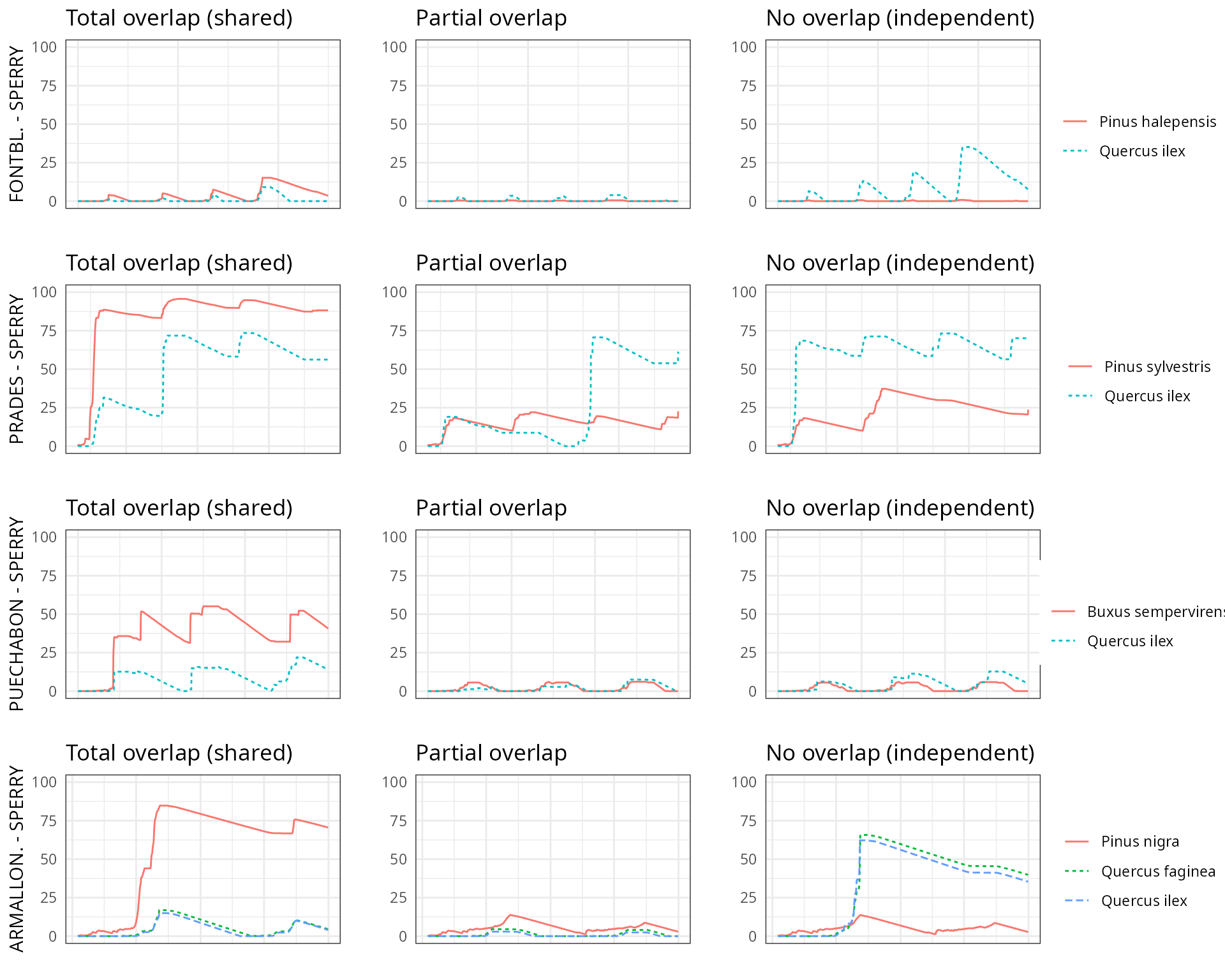
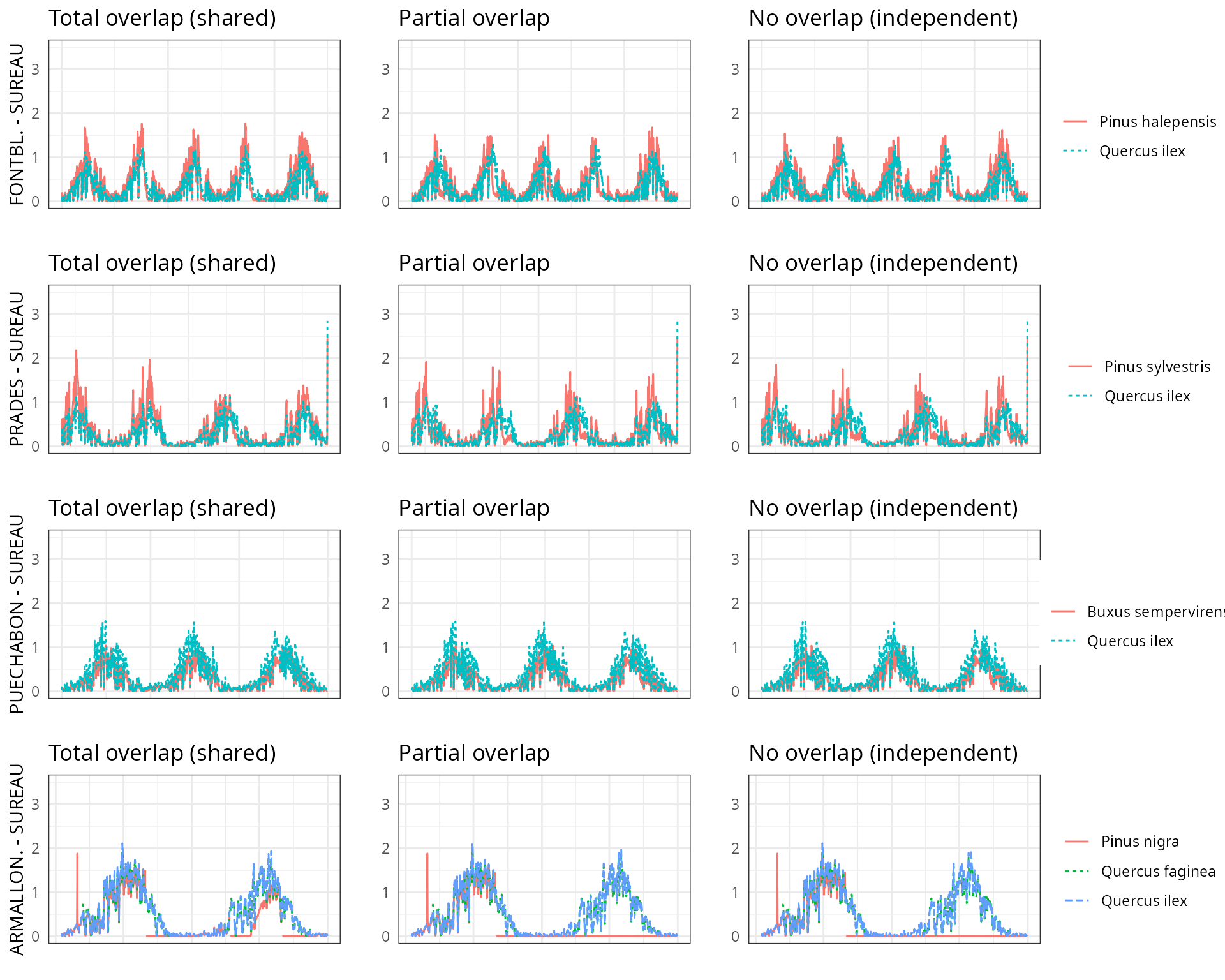
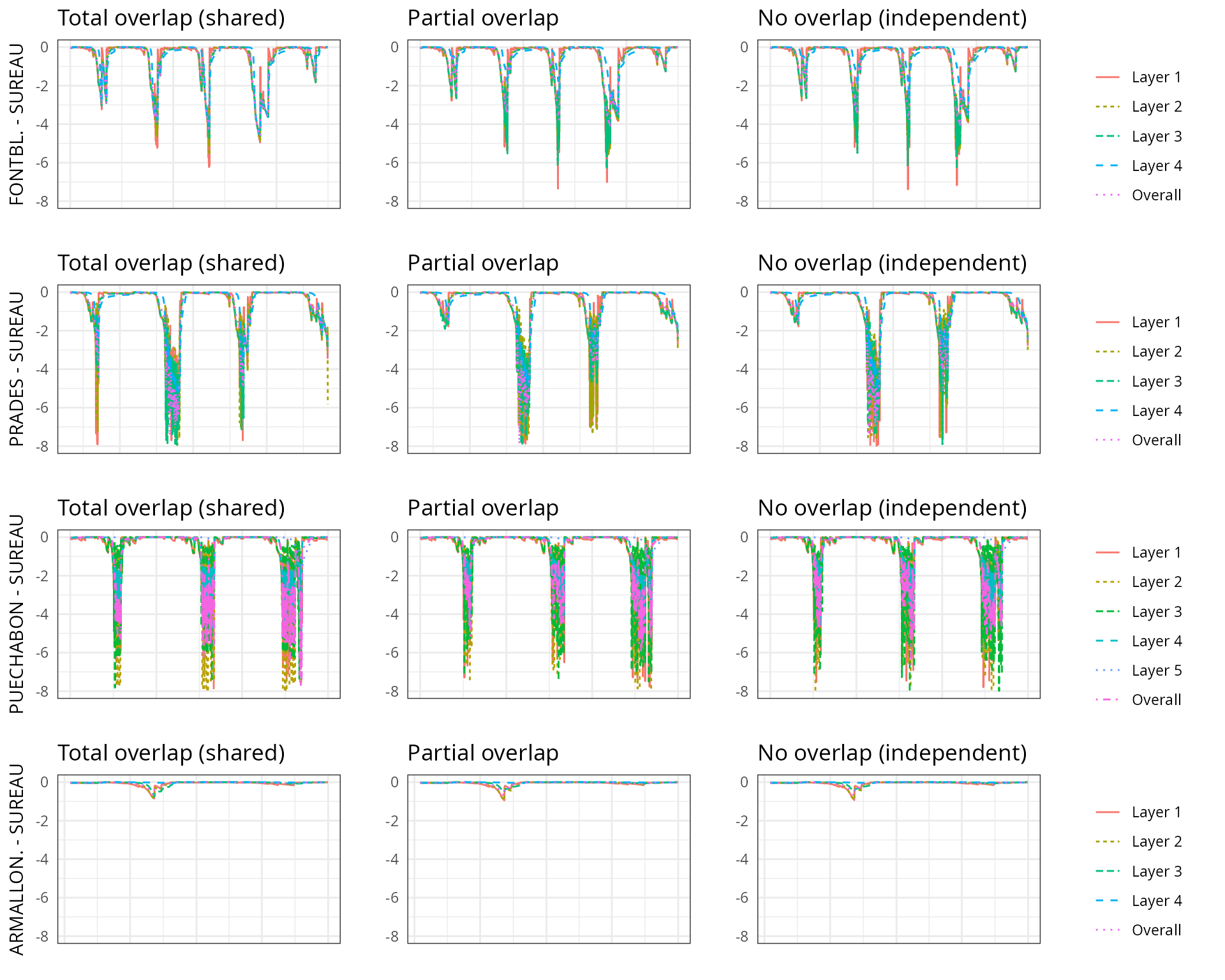
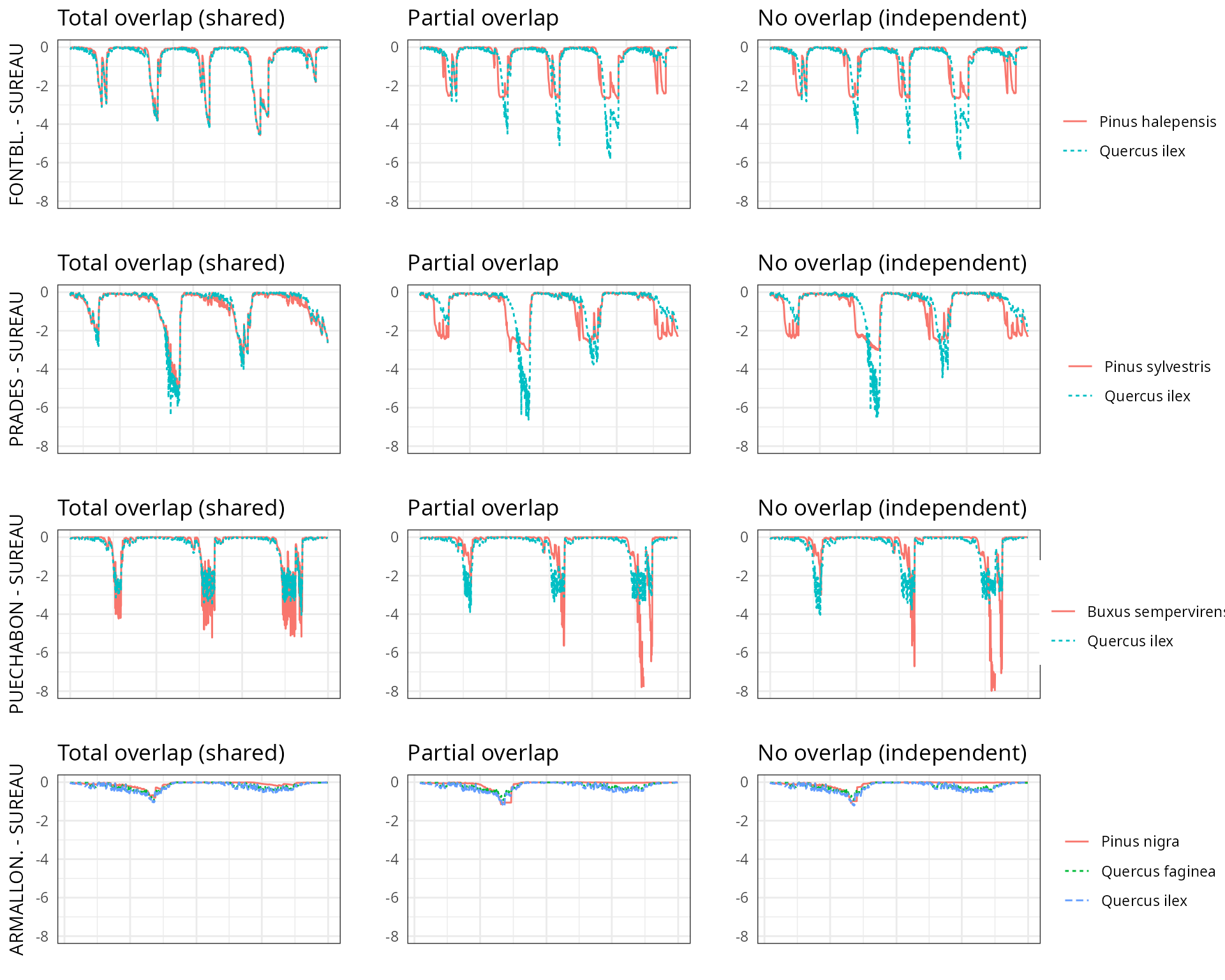
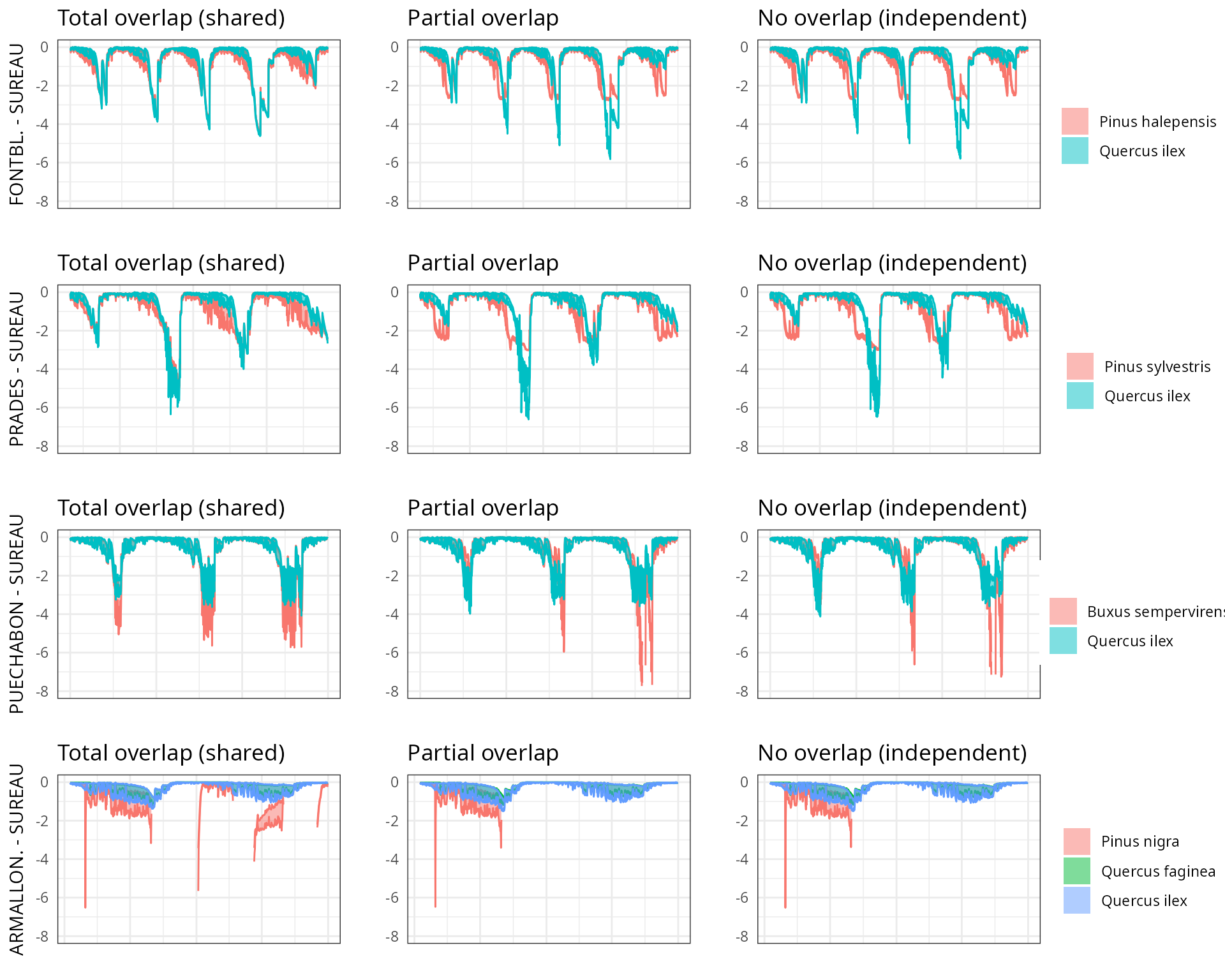
The aim of this vignette is to illustrate the sensitivity of the advanced water balance model (De Cáceres et al. 2021) to the consideration of independent water pools.
Simulations
As target forest stands we take five experimental plots whose data was already used when presenting the water balance model in De Cáceres et al. (2021). An evaluation of model performance in these experimental plots is given in Stand-level evaluation.
For each of these plots we performed simulations using combinations of the three different transpiration modes:
transpirationMode = "Granier"transpirationMode = "Sperry"transpirationMode = "Sureau"
And different assumptions with respect to rhizosphere overlap:
- Default assumption of complete rhizosphere overlap
(
rhizosphereOverlap = "total"). - Assuming partial dynamic rhizosphere overlap
(
rhizosphereOverlap = "partial"). - Assuming complete independence of water pools
(
rhizosphereOverlap = "none").
Recovery from embolism was assumed to depend on soil moisture
(stemCavitationRecovery = "rate" and
leafCavitationRecovery = "rate").

Bibliography
- De Cáceres, M., M. Mencuccini, N. Martin-StPaul, J.-M. Limousin, L. Coll, R. Poyatos, A. Cabon, V. Granda, A. Forner, F. Valladares, and J. Martínez-Vilalta. 2021. Unravelling the effect of species mixing on water use and drought stress in Mediterranean forests: A modelling approach. Agricultural and Forest Meteorology 296:108233.
- Schume, H., G. Jost, and H. Hager. 2004. Soil water depletion and recharge patterns in mixed and pure forest stands of European beech and Norway spruce. Journal of Hydrology 289:258–274.